2021-11-18 22:12


扫码打开虎嗅APP

本文来自微信公众号:神经现实(ID:neureality),作者:Philip Ball,译者:安迪,编辑:Nevaeh,校对:顾金涛,原文标题:《重塑细胞做决定的权利》,题图来自:《异形:契约》
主体能动性,一个被回避的重要问题
在动物免疫系统中,有一种叫做巨噬细胞的白细胞,它们会吞噬并消化异物。这些细胞目标坚定、兴致高昂地追逐它们的猎物:在显微镜下,你可以看到玻片上有团状的巨噬细胞在追逐细菌。细菌试图穿行于路障般的红细胞之间,巨噬细胞则紧随其后改变自己的行进方向。最终,巨噬细胞抓住了逃亡的细菌,将其一口吞下。
请等一下,刚刚的叙述,难道不是在用荒谬的拟人手法来描述一个生物过程吗?单细胞生物并没有自己的心智,所谓的“目标坚定”、“兴致高昂”不该只是人类的臆想吗?你可能会认为这是1944年的一项经典心理学实验的现实版本。
弗里茨·海德(Fritz Heider)和玛丽安·西梅尔(Marianne Simmel)通过那项实验表明,人类倾向于为我们看到的东西赋予目标、编构故事。尽管人们观看的只是抽象几何图形(一个圆形和两个三角形)的移动,但大多数人都会自己编构出颇有戏剧性的追逐与营救的故事。

目的之为错觉?追逐细菌的巨噬细胞,以及海德-西梅尔实验(1944)所用短片的一帧。
不过,我们感觉巨噬细胞有自己的目标,只是出于人类讲故事的倾向吗?毕竟,如果不提及“寻找并吞噬异物”这样的目标,我们甚至没办法描述什么是巨噬细胞。生物学的一大困境就在于如何处理“主体能动性”这种描述,即生物体为了达到特定目的改变环境和自身的能力。生物学领域总是倾向于回避这一问题,一讨论到目标就加上双引号:细胞和细菌并不是真的“试图”做什么,就像生物体的演化并不是“为了”实现什么。
这样一来,一切都归结到基因和分子,物理和化学——不过是无目的事件自然发生罢了,人类却倾向于把它讲成有目标和计划的故事。然而,我们目前有越来越多的理由怀疑,主体能动性并非只是人为编造的故事,而是一种真正的自然现象,甚至不一定为有机体所独有。
了解什么使得一个实体成为主体——改变自身行为和环境来达到特定目的,有助于调和棘手的目的和功能的概念,与生物学之间的矛盾。构建自下而上的主体能动性理论,能够帮助我们理解从细胞到社会的各类事物,甚至还有一些“智能”机器与技术。目前,人们开始怀疑人工智能系统是否会自己发展出主体能动性,但我们甚至不知道主体能动性本身意味着什么。不过,下面提供了一些解决的思路。

- Evangelia Philippidis -
对主体能动性的理解:局限与误区
生物学家恩斯特·迈尔(Ernst Mayr)的著作《什么使得生物学如此特殊?》(What Makes Biology Unique?)是生物学家努力与目的论划清界限的代表。尽管书中承认,生物学领域无法避免从功能的角度谈论问题:比如,眼睛的演化是为了更好地在环境中移动,乳糖酶的功能是分解乳糖,等等……生物学家们不采用这种思维方式就几乎无法做研究。
然而,他们会迅速解释这只是个修辞手法罢了;生物体的行动仅仅由基因所决定,而基因的编码方式又被自然选择所决定。甚至在人文科学领域,主体能动性概念也曾受到抵制。美国心理学家斯金纳在20世纪30年代提出了新行为主义理论,这一理论直到20世纪80年代都是心理学的一个重要分支。
他强调,动物的心智起初就像白板一样,它们的行为只是经由训练所习得的(即“条件反射”)行为。即便是现在,人们也普遍不愿接受其他动物的认知过程代表了真正的自主选择。目前所流行的论调是把所有生物体都当成理查德·道金斯(Richard Dawkins)口中被基因所制造的“机器”。
迈尔认为,生物学这门学科的独特性,体现在它的研究对象拥有一套程序,能够将目的、计划和主体能动性编码到相应的行为。根据这种观点,主体能动性只是被遗传和演化历史所触发的幽灵,即使动物当下的行动看上去存在目的,也不被认为是主体能动性的体现。

- Eric Chow -
但这种观点并未对主体能动性做出解释,只是试图把这个问题消解掉。单个基因并没有主体能动性,所以并不是聚集足够多的基因,就可以产生主体能动性。而这种把主体能动性归因于基因组的看法,既没有告诉我们主体能动性是什么,也没有说清楚是什么使主体能动性得以体现。此外,在任何条件下基因都不能完全决定行为。
基因仅仅表明了一种倾向,即便已经充分了解一个生物体的全部基因和细胞,也无法精确预测其行为。不仅人类如此,结构非常简单的生物也是一样。如果所有的行为都被提前设定好,生物个体就不能对新问题创造出新的解决方法。
而我们知道动物做得到,就像在人为制造的难题前,新喀鸦会就地取材,制造并使用工具来获取食物。这揭示了主体能动性的关键:面对不曾预见的新情况时,做出合适的选择。想象一只野兔被狼所追赶,我们没办法预测它将如何奔跑、转弯,即便狼的行为会随野兔而改变,也无从得知野兔的逃跑策略究竟能否成功。在这个例子中,野兔和狼都运用了主体能动性。或许巨噬细胞所在的认知环境不像狼那样丰富,但很难定义区别到底在哪里。
这个问题之所以令人困惑,可能是因为人们普遍认为复杂的自主行为需要同样复杂的心智来支持。类似植物是否有知觉和意识这一问题,目前就在植物生物学领域掀起了极大的波澜。比如植物会根据过去的经验选择生长方向,而这一过程还可能涉及到电信号,联想到神经元也会互相传递电信号,这些行为都使得植物的活动看起来像是有目的,甚至“有意识”。
但是如果把主体能动性分解一下,就能发现至少传统意义上“会思考”的心智,并非其必要条件。主体能动性可以分解为两部分:一是对相同(或同等)刺激产生不同反应的能力;二是以目标导向的方式做出选择。总的来说,这两项都不是人类所独有的,也并非只有大脑才能做到。

不必依赖复杂心智的主体能动性
第一部分,对给定的刺激做出不同反应的能力,是最容易获得的,只需要类似于抛硬币的行为随机性。这种不可预测性有其演化学上的意义,比如使得捕食者难以预测被捕食者的行动。如果一个生物体对同一个刺激总是做出同样的反应,那它就会成为捕食者的活靶子。
以纤毛虫这种单细胞水生生物为例,当研究者向纤毛虫喷射塑料珠子,以模仿捕食者的侵袭行为时,它或是收缩、或是脱离附着的表面飘走,概率各为50%。显然,没有神经系统也能产生行为随机性。
行为随机性并不等于主体能动性,但前者是后者的必要条件。在随机行为产生的可能选项中,以目标为导向做出选择,这才是真正的主体能动性。英国华威大学的认知科学家托马斯-希尔斯(Thomas Hills)指出,对人类而言,这个选择过程往往需要经由特定的神经环路完成复杂的认知过程。对人类而言,“自由”选择需要我们根据过去的经验,想象未来的可能情景;
要求我们有能力构建环境中的自我形象:一种“丰富的认知表征”,使得这些想象的情景足够精细,能够可靠地预测结果;它也需要抵抗干扰、专注目标的能力。希尔斯说,这并不是人类所独有的,其他动物也能够“选择性地重现与过往经历相关的神经激活模式”,就好像是为未来的类似经历做准备一样。
但这可能需要某种特定的心智“架构”。然而,如果我们想深究细胞或纤毛虫主体能动性的成因,就不能让主体能动性依赖于如此复杂的认知资源。更何况,不仅仅是简单生物体,非生物系统也能表现出了在行为选项中做选择的能力。

主体能动性与信息物理学
为了探究主体能动性的核心,我们需要先把生物学抛在脑后。让我们从信息物理学的角度,看看信息处理过程在引发可变性上的作用。这并不是种新方法。在19世纪中期,物理学家认为,宇宙中的所有变化都受热力学第二定律的制约。该定律指出,变化会导致熵增(通俗点说,就是无序性的增加)。
正是由于热力学第二定律,热量才会自发地从热的区域移动到冷的区域。1867年,物理学家詹姆斯·克拉克·麦克斯韦(James Clerk Maxwell)找到了热力学第二定律的一个“漏洞”。他想象出一种后来被称为麦克斯韦妖的微观存在。假定有一个绝热容器被分成相等的两格,“妖”控制着它们中间的小门,它能选择性地让速度快的粒子进入一格,而让速度慢的粒子进入另一格。这样,热的(速度快的)粒子和冷的(速度慢的)粒子会分别聚集在两格。这违背了热力学第二定律:根据热力学第二定律,热量会发生扩散,直到气体具有均匀的温度。
这个思想实验中的“妖”表现出了主体能动性(尽管现实中麦克斯韦妖是不存在的,因为获取每个粒子的动量这种微观信息是做不到的)。为了实现自己的目标,它选择只为一些特定的气体粒子打开小门。为了试图理解麦克斯韦妖是否真的违背了热力学第二定律,后来的科学家们用一种定义更清晰的假想机械装置代替了麦克斯韦妖,比如用一个电子控制活板门,配上能够感知粒子速度或能量的设备。
这相当于通过预先设计来实现主体能动性:指定了如何利用从环境中收集到的信息来决定行动方案(是否开门)。这样,麦克斯韦妖就成为了一个“信息引擎”,即一种利用信息做功(把气体分成热气和冷气)的机械装置。正是通过这种方式,麦克斯韦妖最终被科学家们驯服了。
在20世纪60年代,物理学家罗夫·兰道尔(Rolf Landauer)表明,麦克斯韦妖终将偿还它在隔离冷热分子时产生的熵损失。为了使用它收集到的粒子运动信息,它必须首先将这些信息记录在某种存储器中。容器中包含了大量气体分子,可是这个存储器的容量有限。因此,存储器必须每隔一段时间就被清空,为新的信息腾出空间。而兰道尔表明,这种清空带来了不可避免的熵开销。所有因冷热分离而损失的熵都会在清空存储器时得到补偿。
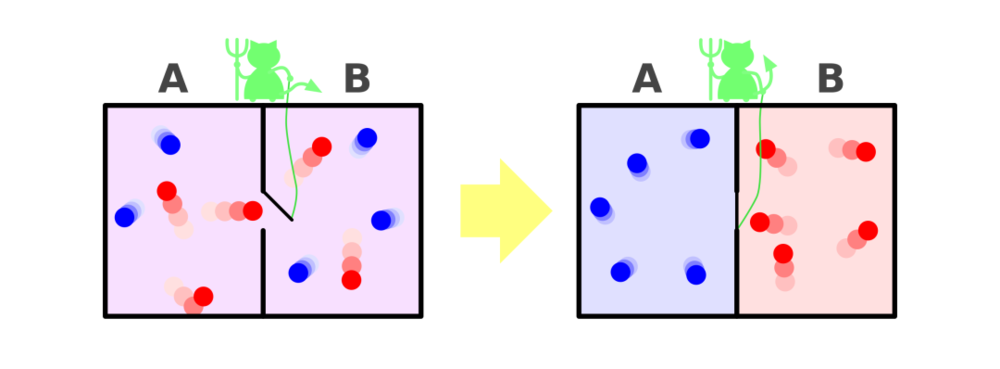
麦克斯韦妖概念图—维基百科
兰道尔的分析揭示了热力学和信息之间的深刻联系。信息,更确切地说,是存储环境信息的能力,是必须不断补充的燃料。麦克斯韦妖所实现的恰恰是物理学家埃尔温·薛定谔(Erwin Schrödinger)在20世纪40年代确定的生物体特征:在热力学第二定律制造混乱和无序的情况下,创造并维持秩序。薛定谔指出,生物体最终也必须通过增加其周围环境的熵来为其内部组织和秩序付出代价——这就是为什么我们的身体会产生热量。
他开创性的著作《生命是什么》(1944)对一批科学家产生了很大的影响,使他们的研究从物理学转向了生命科学,其中就包括弗朗西斯·克里克(Francis Crick),他和詹姆斯-沃森(James Watson)在1953年发现了DNA的双螺旋结构。薛定谔相信生命的主体能动性完全由一套编码指令所维持,这套指令指定了生物体对环境做出怎样的反应,他称之为“非周期性晶体”(aperiodic crystal),后来被发现的DNA就被沃森和克里克认为是这种非周期性晶体,当然,DNA在很大程度上解释了生物体如何运作。
但这一观点没有为真正的主体能动性的偶然性、情境性和多变性留下空间。很可惜,薛定谔未能将“生命摄入负熵”这一看法,与他的物理学家同事们在麦克斯韦妖方面做的工作联系起来。这让生物学界错失了不再将主体能动性视为虚构或幻觉的大好机会。复杂系统理论家斯图尔特·考夫曼(Stuart Kauffman)和哲学家菲利普·克莱顿(Philip Clayton)认为:“想完全理解生物的主体能动性,其必需条件尚未得到阐述:一套坚实的生物体理论。”

关于主体能动性的新研究
随着科学家们对信息论、热力学和生命科学交叉领域的探索,组织、信息和主体能动性之间的关联一点点被揭示出来。2012年,苏珊娜·斯蒂尔(Susanne Still)与位于加州的劳伦斯伯克利国家实验室的加文·克鲁克斯(Gavin Crooks)等人合作,发现了为何对于有目标的主体而言记忆至关重要。有了记忆,主体就能存储环境的表征,利用表征对未来进行预测,并尽可能以最有效率的方式利用能量。
能量效率(energy efficiency)显然是演化生物学中的一个重要目标,浪费的能量较少就意味着获取能量所需的时间较少。热力学表明,虽然任何有效功都要付出熵的代价,做功效率越高,无用的热量散失就越少。
麦克斯韦妖逃不了熵债,因为它的记忆有限,并且会被清除。即便如此,麦克斯韦妖还是能够控制随机粒子的运动,因为它能掌握任何生物体都无法接触到的分子层面的信息。但是,当你不那么全知全能,却想要实现目标的时候,预测能力就很重要了。

- Gracla Lam -
“大多数真实的系统,特别是生物系统,都存在‘感知过滤器’”,斯蒂尔说,“这意味着它们不能接触现实世界的真正状态,而只能测量其中的某些方面。它们被迫依靠仅有的知识运作,并基于部分信息做出推断。”
通过研究真实世界中的“信息引擎”怎样应对这种只能接触到部分信息的情况,斯蒂尔和克鲁克斯等人发现,行为的效率取决于专注力,即主体专注于识别和存储对实现目标有帮助的信息的能力。研究表明,主体在记忆中存储的“无用”信息越多,行动效率就越低。总之,主体想提高效率,就得会鉴别信息。
不过,尽管目前的研究构建了大的框架,还有一些难点有待解决。一般来说,环境并非是静态的,而是受到主体的影响,因此仅仅学习环境本身的信息还不够。斯蒂尔说:“主体会改变它所要学习的过程本身。”这使得情况更加棘手。主体可能面临着适应环境或改变环境的抉择:有时绕过障碍更好,但有时挖地道过去反而更佳。
更重要的是,只有当环境能够对这一策略做出响应时,采取行动才是有效的。如果与周围环境的响应相比,主体试图改变环境的速度更快,反而没什么意义。比如,当你摇盐罐摇得太快(在超声波频率下),并不能把盐倒出来。正如斯蒂尔所说,“在一串特定行为和环境的动态之间可能存在某种‘阻抗匹配’(impedance match)。”
而在现实生活中,主体可能必须在几个相互冲突的目标中找到好的折衷方法。斯蒂尔说,她现在正在积极研究“信息引擎”主体如何应对这些难题。然而,如果主体将要遇到的每一种情况并没有被预先编码过,它如何找到实现目标的方法?
对人类而言,这往往源于思考、经验和直觉,换句话说,源于复杂的认知过程。然而,即使是没有一丁点认知的“最小的主体”,似乎也能找到创造性的策略。2013年,哈佛大学的计算机科学家亚历山大·威斯纳·格罗斯(Alex Wissner-Gross)和现在在麻省理工大学工作的卡梅伦·弗里尔(Cameron Freer)表明,简单的优化规则就可以让简单的无机物体产生非常类似生命体的行为:例如,诱导它们合作完成一项任务,或是将其他物体作为工具。

威斯纳·格罗斯和弗里尔用计算机模拟了二维空间中移动的圆盘,这种移动有点像显微镜载玻片上细胞或细菌的运动。圆盘可以沿着空间中的任意路径移动,但它需要服从一个简单的规则:圆盘的移动和与其他圆盘的互动需要能够最大化它在一段时间内的熵。
粗略地讲,这个熵的最大化相当于为物体的移动方式保留了尽量多的可能性,比如说,它可以留在开阔的区域,避免被困在狭窄的空间中。这种规则就像一种引导物体移动的力,他们将其命名为“熵力”。奇怪的是,由这种规则产生的行为看起来像是目标导向且具有智能的行为。
在一个例子中,一个大圆盘“利用”一个小圆盘从狭窄的管子中取出另一个小圆盘,看起来就像在使用工具一样。在另一个例子中,不同隔间中的两个圆盘同步运动,把另一个较大的圆盘移动到它们可以与之互动的位置,看起来很像社会合作行为。
对这种场景的计算机模拟可能会让我们想到之前提到的海德-西梅尔实验。但后者涉及到人为设计的动作,前者则只基于系统的总熵随时间变化的模型。主体设想它们的行为发生在未来,计算出行为对总熵的影响,做出自己的“选择”。

- Julia Yellow -
在现实世界中,自主进行这类计算的物体需要为当前环境中可能的运动轨迹构建内部表征,这可以说是一种工作记忆。它们还需要计算资源来计算行为引发的结果。不过威斯纳·格罗斯和弗里尔的模型并不是针对生物学上的实际情况搭建的。
相反,关键就在于,解决复杂问题的主体能动性可以从简单的熵规则中出现。这并不是说生物体的主体能动性真的以这种方式产生,而是说,简单的物理规则,而非复杂的认知过程,就足以产生复杂的、目标导向的行为。
对主体能动性的新表述
因此,我们不必借助神秘主义也可以描述主体能动性怎样产生。演化的过程创造并强化了能量效率等目标,但并未指定主体实现目标的方式。而一个试图提高能量效率的主体会演化出一种记忆,以储存并表征环境中对实现目标有用的部分。这便是产生主体能动性的原材料。与此同时,生物体将演化出一种能力,在应对基本相同的刺激时产生多种可选的行动方案。
起初,在方案间的选择可能是随机的。但是,具有记忆的生物体,能够“思考”可能的选项,基于对环境的内部表征做出更有效的选择。这并不一定需要大脑(尽管大脑肯定能帮的上忙)。简而言之,这就是主体能动性。

- Eiko Ojala -
事实当然未必如此。但是这种描述的优点在于把复杂的主体能动性分解成更简单的过程,而不依赖于高度专门化的(甚至不依赖于“有机的”)硬件基础(比如大脑)。它也反映出,复杂的认知能力似乎常常是从为了其他目的而产生的能力的融合中演化而来的。
至少,这项最新研究表明,主体能动性绝不仅仅是演化的古怪副产品,也不是人类的拟人化倾向导致的错觉。相反,主体能动性似乎是物质的一种偶然却非凡的属性,当我们想为观察到的事物提供因果解释时,应该放心地援引它。
比如,如果我们想解释为什么火山岩在某个特定位置,只用力学知识就可以,不需要在解释中包括任何目标。地球深处的热量——即分子的运动——和重力共同作用,产生对流,再将岩浆带到地表。但如果我们想解释为什么鸟巢在某个特定位置,如果只是自下而上地解释,不引入鸟为什么筑巢,不谈到鸟的主体能动性,解释就不完整。主体能动性是涌现特性的典型例子之一,它来源于更“基本”的组成成分的属性,却不能被这些属性完全解释。

一个真正的主体能动性理论可能最终有助于澄清科学对自由意志的看法。因为自由意志不过就是主体能动性加上意识:我们能够有目的地做出影响环境的决定,以及意识到自己的行为。从这个观点来看,自由意志的问题没有给意识问题增加新的障碍。
此外,把主体能动性引入其中使得我们能从演化和神经科学的角度,而非哲学里决定论的角度,讨论自由意志的问题。这一切的关键在于,主体能动性,就像意识以及生命本身一样,不是只在细节层面就能理解的。
它也不是粒子运动的附属品,并不是说只要聚集的粒子足够多,它们就表现得“好像”有主体能动性,甚至“好像”有意识。主体能动性是真实存在的,不必以双引号的形式对其降格。对于这一点,等我们找到足够好的理论来解释主体能动性怎样出现时,可能就不会有那么多的反对声音了。
原文:https://aeon.co/essays/the-biological-research-putting-purpose-back-into-life
本文来自微信公众号:神经现实(ID:neureality),作者:Philip Ball,译者:安迪,编辑:Nevaeh,校对:顾金涛
 34:52
34:52








 07:08
07:08
 23:16
23:16
 01:03:10
01:03:10
 05:50
05:50
 07:34
07:34
 11:28
11:28
 12:58
12:58
 27:04
27:04
 20:38
20:38




